Prokaryotische Chromosomen: Nützliche Hinweise zu prokaryotischen Chromosomen
Hier sind Ihre Notizen zu prokaryotischen Chromosomen!
Die genetische Information einer prokaryotischen Zelle wird im Nukleoplasma auf der als bakterielles Chromosom bezeichneten Struktur übertragen. Es besteht aus einem doppelhelicalen DNA-Molekül (Desoxyribonukleinsäure), das niemals mit basischen Proteinen assoziiert ist. In einigen Prokaryoten wurde gezeigt, dass es kreisförmig ist.

Bild mit freundlicher Genehmigung: hangthebankers.com/wp-content/chromosome.jpg
Das bakterielle Chromosom ist folglich nicht strukturell homolog mit den Kernchromosomen der eukaryontischen Zelle, sondern mit der in den eukaryotischen Mitochondrien und Chloroplasten vorhandenen organellaren DNA. Es ist wahrscheinlich, dass ein einzelnes bakterielles Chromosom (dh ein sehr langes DNA-Molekül) alle genetischen Informationen enthält, die erforderlich sind, um die wesentlichen Eigenschaften der prokaryotischen Zelle festzulegen.
Plasmide: 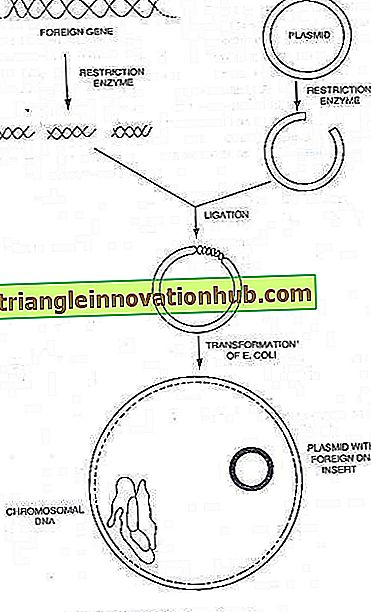

Viele Bakterien können auch kleine, extra chromosomale zirkuläre DNA-Moleküle beherbergen, die zur autonomen Replikation befähigt sind. Diese so genannten Plasmide tragen die Determinanten für solche phänotypischen Merkmale wie Resistenzen gegen Medikamente und andere antibakterielle Substanzen sowie für die Enzyme, die bestimmte Stoffwechselwege vermitteln . Die DNA-Menge in einem Plasmid beträgt 0, 1 bis 5 Prozent der Menge im Bakterienchromosom.
In Escherichia coli wurde festgestellt, dass es zwei Paarungstypen gibt. Während der Konjugation fungiert ein Partner nur als genetischer Spender oder männlich und der andere nur als genetischer Empfänger oder weiblicher Partner. Da die einzige Funktion des Mannes darin besteht, etwas von seiner DNA zu übertragen, muss diese nicht lebensfähig bleiben, während die weibliche Zelle lebensfähig bleiben muss, damit sich die Zygote darin entwickeln kann.
Dies wurde entdeckt, dass die Männlichkeit in Bakterien durch ein übertragbares genetisches Element bestimmt wird, wenn männliche und weibliche Bakterien konjugieren, wobei jede weibliche Zelle in eine männliche umgewandelt wird. Das genetische Element, das die vererbte Eigenschaft der Männlichkeit bestimmt, wird als F-Faktor (F = Fertilität) bezeichnet. Es wird nur durch direkten Kontakt von Zelle zu Zelle übertragen. Lederberg prägte 1952 den Begriff Plasmid als genetischen Namen für alle extra chromosomalen erblichen Determinanten, von denen F ein Beispiel ist.
Es ist nun bekannt, dass Bakterienplasmide kleine, zirkuläre DNA-Moleküle sind, die die Gene für ihre eigene Replikation tragen. In vielen Fällen tragen sie auch Gene, die der Wirtszelle neue Eigenschaften verleihen, z. B. Resistenz gegen Medikamente oder die Produktion von Toxinen. Viele Plasmide tragen Gene, die den Konjugationsprozess steuern. Somit ist die Konjugation ein Mechanismus, der der Bakterienzelle durch ein Plasmid auferlegt wird, dessen normales Ergebnis der Transfer von Plasmid-DNA ist.
Arten von Plasmiden:
Die meisten Plasmide wurden auf der Grundlage der Wirteigenschaften klassifiziert. So gibt es den R-Faktor (R = Resistenz) und die COI-Faktoren (COI = Colicinogeny) der gramnegativen Bakterien, die Penicillinase-Plasmide von Staphylococcus aureus, die Abbauplasmide von Pseudomonas, die kryptischen Plasmide usw.
Eigenschaften von Plasmiden:
Die Struktur aller bekannten Plasmide besteht aus kreisförmigen doppelsträngigen DNA-Molekülen. Einige von ihnen haben Molekulargewichte im Bereich von 5 × 10 bis 7 × 10. Einer (ein R-Faktor) hat ein Molekulargewicht von nur 1 × 10 7, und einige der kryptischen Plasmide sind sogar noch kleiner. Da die Menge an DNA, die erforderlich ist, um ein durchschnittliches Polypeptid mit einem Molekulargewicht von 40.000 zu codieren, etwa 6 × 10 7 beträgt, kann F 1 und andere Plasmide ähnlicher Größe bis zu 100 Gene enthalten.
Genexpression in Prokaryoten:
Bakterienchromosomen:
Es wurde nun festgestellt, dass im Zentrum der Bakterienzelle immer ein Bakterienchromosom vorhanden ist, das nicht mitotisch und meiotisch zerfallen kann und nicht in einer bestimmten Kernmembran eingeschlossen ist. Eine durchschnittliche Bakterienzelle enthält ein Tausendstel des DNA-Gehalts einer typischen Eukaryontenzelle.
Das bakterielle Chromosom ist ein dünnes, langgestrecktes, flexibles und kreisförmiges DNA-Molekül mit wenigen Proteinen. Die Bakterienzellen können sich sehr schnell teilen. Die Zellteilung wird durch die Verdoppelung aller Zellbestandteile abgeschlossen, gefolgt von der Aufteilung der Zelle in zwei Tochterzellen. Der erste Schritt bei der Zellteilung ist die DNA-Duplikation.
Genetische Rekombination in Bakterien:
Dies ist ein Prozess, bei dem genetisches Material, das in zwei separaten Genomen enthalten ist, in einer Einheit zusammengefasst wird. In Bakterien erfolgt die Rekombination durch (1) Transformation, (2) Transduktion und (3) Konjugation. Diese Phänomene wurden bereits in den vorangegangenen Abschnitten unter der Überschrift „sexuelle Fortpflanzung in Bakterien“ ausführlich erörtert.

Induktion und Unterdrückung:
Das genetische Potenzial eines Virus ist begrenzt und es nutzt im Wesentlichen die Wirtsmaschinerie für seine Replikation. Auf der anderen Seite können die Bakterien ihre Bestandteile aus einfachen Salzen und Zucker herstellen. Das Bakterium Escherichia coli hat etwa 2500 Gene auf seinem Chromosom. Das Bakterium behält seine innere Ökonomie nur bei, indem es die Wirkung von wenigen Genen gleichzeitig erlaubt.
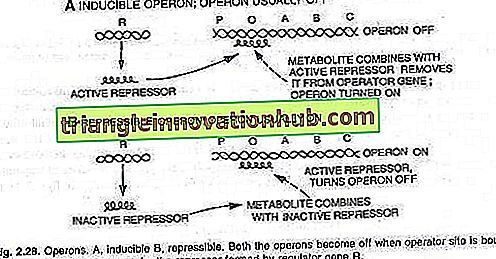
Dies bedeutet, dass Gene je nach Anforderung ein- oder ausgeschaltet werden. Ein Satz von Genen wird eingeschaltet, wenn ein neues Substrat gehandhabt und metabolisiert werden muss. Wenn der Satz dieser Gene eingeschaltet ist, werden Enzyme produziert, die das neue Substrat metabolisieren. Dieses Phänomen wird als Induktion bezeichnet und die Moleküle (Metaboliten), die dieses Phänomen hervorrufen, sind Induktoren.
Wenn andererseits ein von dem Bakterium benötigter Metabolit im Übermaß von außen zugeführt wird, hemmt das Bakterium die Bildung, wodurch die weitere Produktion des Metaboliten durch das Bakterium gestoppt wird. Diese inaktivierten Gene werden daher als unterdrückbar bezeichnet, und das Phänomen wird als Repression oder Feedback-Repression bezeichnet.
Einige der Gene werden jedoch ständig exprimiert, um die normale Zellaktivität wie die Glykolyse zu gewährleisten. Diese Gene sind als konstitutiv bekannt, und die von ihnen produzierten Enzyme sind als konstitutive Enzyme wie Dehydrogenasen bekannt.
Operon-Konzept:
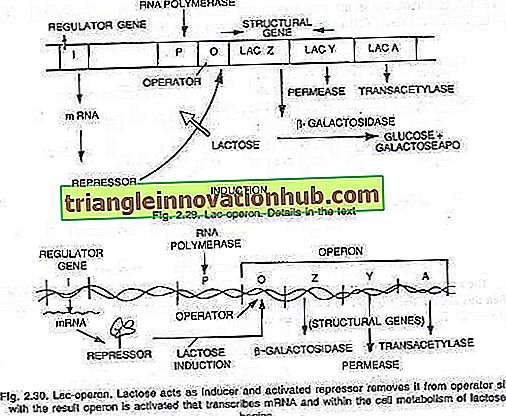
Francois Jacob und Jacques Monod (1961), während sie am Pasteur-Institut in Paris arbeiteten, schlugen vor, dass Stoffwechselwege als Einheit reguliert werden. Sie untersuchten die Veränderung der Art von Enzymen von E. coli, wenn sie mit verschiedenen Zuckersorten versorgt wurden. Sie fanden heraus, dass, wenn die Zuckerlaktose zu den Kulturen von E. coli gegeben wird, drei Enzyme induziert werden, die zum Abbau der Laktose in Glukose und Galaktose erforderlich sind.
Diese neu synthetisierten Enzyme sind (i) p-Galactosidase, (ii) Permease und (iii) Transacetylase. Demnach wird die Synthese dieser drei Enzyme durch ein langes DNA-Segment gesteuert, das als Operon bekannt ist und in eine Operatorstelle O und drei Strukturgene Z, Y und A unterteilt werden kann. Die Wirkung von Strukturgenen wird durch die Operatorstelle mit der reguliert Hilfe eines Repressorproteins, das durch die Wirkung des Gens 'i', das als Regulatorgen bekannt ist, produziert wird.
Die Gene werden ausgedrückt oder nicht ausgedrückt, je nachdem, ob der Bedienerschalter ein- oder ausgeschaltet ist. Wenn der Schalter eingeschaltet ist, werden die drei Gene von RNA-Polymerase in einen einzigen Abschnitt der Messenger-RNA transkribiert, der alle drei Gene abdeckt. Jedes Strukturgen ist allgemein als Cistron bekannt und die transkribierte lange mRNA, die alle Cistrons abdeckt, ist als Polycistron bekannt.
Das Ein- und Ausschalten des Bedienerschalters wird durch ein als Repressor bezeichnetes Protein erreicht. Wenn dieses Protein mit dem Operator verbunden ist (O) und es blockiert, wird der Schalter ausgeschaltet und die drei Gene (Z, Y, A) werden nicht exprimiert (siehe Abb. 2.28).
Lac-Operon:
Dies ist ein Beispiel eines induzierbaren Operons. Durch die Wirkung der Enzympermease werden einige Moleküle Lactose in die Zelle gegeben, von denen eine geringe Menge auch unter verdrängten Bedingungen gefunden wird. Diese wenigen Moleküle werden dann in eine aktive Form von Laktose umgewandelt, die an den Repressor (das Genprodukt) bindet, und der Repressor kann sich daher nicht mehr an den Operator-Switch binden.
Wenn der Operator frei von Repressor-RNA-Polymerase ist, beginnt die Transkription des Operons durch Binden der Promotorstelle P. Die Messenger-RNA, die allen drei Enzymen entspricht, wird synthetisiert, wodurch sie translatiert wird, um drei Enzyme, P-Galactosidase, Permease und Transacetylase zu erzeugen.
Mit der Produktion dieser drei Enzyme beginnt der Metabolismus von Laktose. Die Synthese von Enzymen wird fortgesetzt, sofern und bis alle Laktosemoleküle von der Zelle verbraucht sind. Wenn die letzten an den Repressor gebundenen Laktosemoleküle ebenfalls verbraucht werden, wird der inaktive Repressor aktiv und bindet an die Operator-Site (O), um das Operon normal abzuschalten (siehe Abb. 2.29 und 2.30).
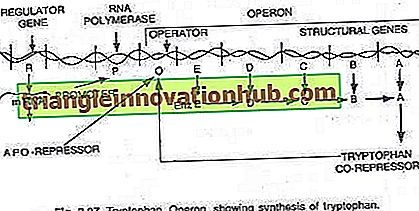
Tryptophan-Operon:
In diesem Fall ist das Operon im Allgemeinen eingeschaltet, so dass Transkription und Translation zur Synthese von Enzymen erforderlich sind, die für die Synthese eines bestimmten Metaboliten durch die Zelle benötigt werden. Das Operon kann jedoch ausgeschaltet werden, wenn die Zelle keinen Metaboliten benötigt oder der Metabolit im Überschuss produziert wurde. Das Tryptophan-Operon besteht aus fünf Genen (trp E, D, C, B und A), die für fünf Enzyme (En z E, D, C, B und A) kodieren, die die Synthese von Tryptophan (einer Aminosäure) katalysieren und somit konstituieren eine anabole Bahn. Die Anwesenheit von Tryptophan dient dazu, die Synthese der für seine Herstellung verantwortlichen Enzyme zu unterdrücken (siehe Abb. 2.27 und 2.31).
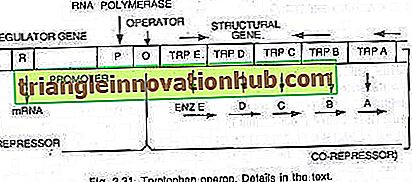
Die fünf Strukturgene, die für diese fünf Enzyme kodieren, werden durch einen einzigen Bedienerschalter reguliert. Hier bindet das von den regulatorischen Genen produzierte Repressorprotein nicht selbst an den Operator. Wenn jedoch Tryptophan hinzugefügt wird, bindet es an den Repressor und wird als Co-Repressor bezeichnet. Der Repressor-Corepressor-Komplex bindet an den Operator und blockiert den Ausdruck dieses Operons.
Die Struktur des Tryptophan-Operons ist der Struktur des Lac-Operons mehr oder weniger ähnlich, weist jedoch eine funktionelle Variation auf. In diesem Fall entspricht das R-Genprodukt dem I-Gen von lac-Operon. Das R-Genprodukt produziert Protein, das an sich nicht an den Operator binden kann. Dies wird als Apo-Repressor bezeichnet.
In Gegenwart von Tryptophan, einem Co-Repressor, wird der funktionelle Repressor gebildet, der sich nun an den Operator bindet und die Transkription des Operons und die Produktion von Tryptophan verhindert. Hier erklärt das Operon-Modell sowohl das Induktions- als auch das Repressionsphänomen in Prokaryoten.


