5 Hauptfunktionen der Antikörper von B-Zellen
Einige der Hauptfunktionen, die das ausgeschiedene Immunglobulin oder die Antikörper von b-Zellen ausführen, lauten wie folgt:
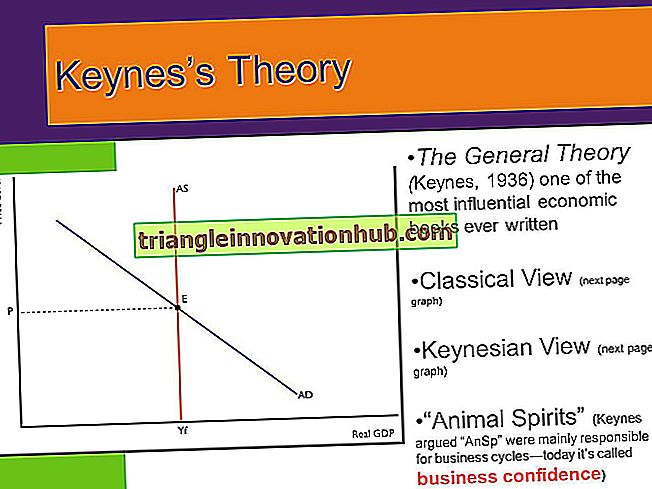
Funktionen von Antikörpern:
Antikörper an sich können das Antigen nicht abtöten und / oder entfernen. Der Zweck des Antikörpers besteht darin, an das spezifische Antigen zu binden.
Infolge der Antikörperbindung werden viele Ereignisse durch die Fc-Region initiiert, die für die Beseitigung von Antigen verantwortlich sind:
1. Gegen eine Mikrobe induzierter Antikörper bindet an die Mikrobe durch die Fab-Regionen des Antikörpers.
Bei der Bindung des Antikörpers an die Mikrobe wird der klassische Komplementweg durch die Fc-Region des Antikörpers aktiviert. Die klassische Aktivierung des Komplementweges führt zur Bildung von Poren in der Zellwand der Mikrobe, was zum Absterben der Mikrobe führt. Bakterien und Viren in der extrazellulären Umgebung werden durch die Komponenten des klassischen Komplementweges abgetötet.
2. Die Zellmembranen einiger Effektorzellen (wie Makrophagen und NK-Zellen) haben Rezeptoren für die Fc-Region der Antikörper.
Der Fc-Rezeptor an der Makrophagenmembran bindet an die Fc-Region des Antikörpers im Antigen-Antikörper-Komplex (Abbildung 9.8). Somit ist das Antigen indirekt durch den Antikörper mit dem Makrophagen verbunden.
Danach macht der Makrophagen den Antigen-Antikörper-Komplex opson. (Opsonisierung ist die Förderung der Phagozytose von Antigenen durch Phagozyten. Mit dem Begriff Opsonin wird eine Substanz beschrieben, die die Phagozytose verstärkt. Antikörper und Komplementkomponenten C3b und C4b sind Opsonine.) Die verschlungenen Mikroben werden durch die lysosomalen Enzyme der Makrophagen abgetötet.
3. Antikörperabhängige zellvermittelte Zytotoxizität (ADCC):
Abgesehen von der Abtötung von Bakterien und Viren können die Antikörper auch die Zerstörung anderer Zellen vermitteln, die Antigene auf ihren Zellmembranen exprimieren.

Abb. 9.8A bis E:
Die opsonische Funktion von Immunglobulin. (A) Das Antigen bindet an die Fab-Regionen des Antikörpers und bildet den Antigen-Antikörper-Komplex. (B) Die Fc-Region des Antigen-gebundenen Antikörpers bindet an den Fc-Rezeptor auf der Effektorzelle (wie etwa Makrophagen). (C und D) Die Makrophagenpseudopodien umgeben den Antigen-Antikörper-Komplex und (E) Der Antigen-Antikörper-Komplex wird in den Makrophagen eingeschlossen. Der Antigen-Antikörper-Komplex liegt innerhalb eines Membranvesikels im Makrophagen-Zytoplasma

Abb. 9.9 A und B: Antikörperabhängige zellvermittelte Zytotoxizität (ADCC).
(A) Antikörper bindet an das Antigen auf der Zellmembran durch seine Fab-Enden, (B) Antigengebundener Antikörper bindet an den Fc-Rezeptor auf Effektorzellen (wie Makrophagen, NK-Zellen) über die Fc-Region. Die Bindung des Antikörpers an den Fc-Rezeptor aktiviert die Effektorzelle, was zur Degranulation oder Sekretion des Inhaltes der Effektorzelle über dem Antigen führt, was zur Lyse der Zelle führt
Die Fab-Region des Antikörpers bindet an das Antigen, das auf der Zellmembran der Zielzelle exprimiert wird (9.9).
↓
Die Fc-Region des Antikörpers bindet an den Fc-Rezeptor auf Effektorzellen (wie Makrophagen und NK-Zellen).
↓
Die Bindung der Fc-Region an den Fc-Rezeptor sendet ein Signal in die Effektorzelle, was zur Sekretion von lytischen Enzymen und toxischen Substanzen durch die Effektorzelle führt. Die abgesonderten Substanzen töten die Zielzelle.
Die Makrophagen, NK-Zellen, Eosinophilen und Neutrophilen sind in der Lage, an die Fc-Region des Antikörpers zu binden und ADCC zu vermitteln, um Zielzellen zu zerstören. Der ADCC-Mechanismus wird verwendet, um Zielzellen wie mit Krebszellen infizierte Zellen und Parasiten zu eliminieren.
4. Neutralisierung von Toxinen durch Antikörper:
Die durch bestimmte Bakterien produzierten Toxine verursachen Krankheiten (Diphtherietoxin, das durch Corynebacterium diphtheriae produziert wird, verursacht Diphtherie; Tetanustoxin, das durch Clostridium tetani Tetanus produziert wird). Bei diesen Krankheiten sollten die Toxine neutralisiert werden, um den Tod von Patienten zu verhindern. Der Patient wird mit Antiserum (normalerweise bei Pferden gezüchtet) behandelt, das Antikörper gegen das Toxin enthält. Die Antitoxin-Antikörper (im Antiserum) binden an die Toxinmoleküle und neutralisieren die toxischen Wirkungen des Toxins und retten den Patienten.

Abb. 9.1 OA bis C:
Antitoxin-Antikörper binden an Toxinmoleküle und verhindern die Wirkung von Toxin auf die Wirtszelle. (A) Normalerweise binden die Toxinmoleküle an spezifische Toxinrezeptoren auf der Wirtszellmembran. Die Toxinmoleküle werden in die Zelle aufgenommen, wo das Toxin seine Wirkung auf die Funktionen der Wirtszellen ausübt. (B) Antitoxin-Antikörper binden an Toxinmoleküle im Kreislauf und bilden Toxin-Antitoxin-Antikörper-Komplexe.
Die Bildung von Toxin-Antitoxin-Komplex stört die Bindung von Toxin an seine spezifischen Rezeptoren auf der Zelloberfläche. (C) Der Toxin-gebundene Antikörper bindet über die Fc-Region des Antikörpers an den Fc-Rezeptor auf der Makrophagenmembran. Folglich wird der Toxin-Antitoxin-Antikörperkomplex vom Makrophagen verschlungen und anschließend zerstört
Normalerweise bindet das Toxinmolekül an spezifische Toxinrezeptoren auf Zellen. Das Toxin wird in die Zelle aufgenommen, wo das Toxin seine toxische Wirkung entfaltet. Um toxische Wirkungen zu erzeugen, sollte das Toxin daher in den spezifischen Zelltyp eingehen. Die Antitoxin-Antikörper im Antiserum binden an die Toxinmoleküle und verhindern die Bindung von Toxin an spezifische Toxinrezeptoren.
Die Fc-Regionen von Toxin-gebundenen Antitoxin-Antikörpern binden an Fc-Rezeptoren in Makrophagen und führen zur Einengung des Toxin-Antitoxin-Antikörperkomplexes durch die Makrophagen.
5. Verhinderung einer Infektion durch Sekretions-IgA auf Schleimhautoberflächen:
IgA-Antikörper, die in den Schleimhautsekreten (des Gastrointestinaltrakts, des Urogenitaltrakts und der Atemwege) vorhanden sind, binden an bakterielle Flagellen und können die Beweglichkeit der Bakterien beeinträchtigen. (Flagellen sind die Beweglichkeitsorgane von Bakterien.) Infolgedessen werden die Chancen beeinträchtigt, dass Bakterien durch die Schleimhaut in den Wirt gelangen.
IgA-Antikörper in Sekreten können auch an Adhäsionsmoleküle auf der bakteriellen Oberfläche binden und dadurch die Adhäsion von Bakterien an das Schleimhautepithel des Wirts beeinträchtigen; und folglich wird der Eintritt von Bakterien in den Wirt gestört. IgA-Antikörper in den Darmsekreten binden an spezifische Viruspartikel und verhindern die Virusinfektion.
IgM- und IgG-Klassen von Antikörpern und Diagnose mikrobieller Infektionen:
Die Immunantwort, die während des ersten Antigeneintritts in den Wirt induziert wird, wird als primäre Immunantwort bezeichnet. Die ruhende B-Zelle teilt sich bei Aktivierung, um Plasmazellen und Speicher-B-Zellen herzustellen. Die von der Plasmazelle (aus einer aktivierten ruhenden B-Zelle stammenden) sezernierten Antikörper gehören immer zur IgM-Klasse.
Die gegen eine Mikrobe gebildete IgM-Klasse von Antikörpern bleibt einige Monate im Kreislauf, und dann sinkt der IgM-Spiegel (Abb. 9.11). Daher zeigt der Nachweis der IgM-Klasse von Antikörpern gegen eine Mikrobe, dass der Wirt kürzlich infiziert wurde. Daher suchen viele immundiagnostische Systeme nach IgM-Antikörpern zur Diagnose einer vorliegenden oder kürzlich durchgeführten mikrobiellen Infektion.

Abb. 9.11:
IgM- und IgG-Klassen der Antikörperproduktion während einer primären Immunantwort. Beim ersten Eintritt des Antigens in den Wirt werden ruhende B-Zellen gegen die Mikrobe aktiviert. Die aktivierten B-Zellen teilen sich, um Effektorzellen (Plasma) und Speicher-B-Zellen herzustellen.
Die Plasmazellen sezernieren Antikörper gegen das Antigen, das für seine Produktion verantwortlich war (durch Aktivierung der ruhenden B-Zellen). Die anfängliche Klasse von Antikörpern, die während einer primären Immunantwort produziert werden, gehört zur IgM-Klasse.
Die IgM-Klasse von Antikörpern bleibt einige Monate im Kreislauf, und dann sinkt der IgM-Antikörper-Spiegel. Wenig später als bei der Herstellung von IgM-Antikörpern werden IgG-Antikörper gegen das Antigen hergestellt. Die IgG-Antikörper bleiben im Vergleich zu IgM-Antikörpern normalerweise länger im Kreislauf. Sowohl IgM- als auch IgG-Antikörper binden an dasselbe Antigen
Während einer primären Immunantwort erscheint die IgG-Klasse von Antikörpern (gegen dasselbe Antigen) wenig später als das Auftreten der IgM-Klasse von Antikörpern. Die IgG-Klasse von Antikörpern bleibt jedoch nach dem ersten Eintritt des Antigens viele Monate im Blutkreislauf (Abbildung 9.11).
Darüber hinaus werden IgG-Antikörper während nachfolgender Infektionen durch dasselbe Antigen in großen Mengen gegen das Antigen produziert. Daher legt das Vorhandensein der IgG-Klasse von Antikörpern gegen eine Mikrobe nur nahe, dass die Person durch die Mikrobe infiziert ist; Wir können jedoch nicht sagen, ob es sich bei der Infektion um eine fortlaufende Infektion oder eine kürzlich durchgeführte Infektion oder um eine Infektion der Vergangenheit handelt, die möglicherweise schon vor langer Zeit aufgetreten ist.